Update.--Elements of metacommunity structure can now be analyzed in R! Tad Dallas has developed an R packed entitled "metacom" that mirrors most of the options available in the Matlab functions.
Conducting analyses.--In conjunction with Christopher Higgins, we have expanded on the original methods of Leibold and Mikkelson to 1) incorporate additional null model options for analyses of coherence, 2) allow analyses beyond the first ordination axis, 3) provide alternatives to reciprocal averaging as an ordination method, and 4) to allow the ordination of axes based on abundances and not just presence-absence data. There are now 10 null model choices for analyses of coherence that allow for greater flexibility depending on the nature of the metacommunity of interest. In addition, because more than one ordination axis may evince ecologically meaningful patterns (Presley et al. 2009, 2011), analyses may be conducted for as many axes as necessary. Besides reciprocal averaging, one may choose to maximally pack a matrix, as for analyses of nested subsets, or to manually define the gradient of analysis. For example, if one is has data along an elevational gradient, metacommunity along the gradient per se may be of interest; therefore, one would not wish to re-order sites or species. The complete MATLAB libraries are available here. Use the function 'metacommunity' to conduct analyses (and the "help metacommunity" command to get get the instructions on how to use the function). These functions are modified and updated from time to time to add flexibility.
We have had a number of requests for assistance in interpreting the output of the functions. This short MS Word document goes step-by-step through an example. If you have any questions on running the analyses, interpreting results, or if you have suggestions for improvement of the approach, feel free to contact me via email.
In addition to the basic "metacommunity" function that runs the procedure to analyze coherence, turnover, and boundary clumping. Chris Higgins has written a function that allows abundances to be used in the ordination of observed matrices. This function is called "metacommunity2". Because the order of sites and species using this approach is sensitive to abundances, it would only be appropriate to use this option when reliable estimates of abundance exist for each species at each site. Otherwise, noise associated with abundance estimation (which is greater than for binary data) may obscure or distort patterns. To date, we have used this approach only for exploratory purposes. NOTE: The current version of the function only uses abundances to order the empirical matrix and not the associated null matrices, soon an update will be available that randomizes populations so that ordered null matrices are more comparable to the empirical matrix under consideration.
An additional auxiliary function exists called mcvi (which stands for metacommunity visual inspection) that will quickly provide you with the ordinated empirical matrix, a matrix of embedded absences for that matrix, and a list of boundaries occurring at each site. This function is useful for quick creation of figures to visualize empirical structures.
|
| Flowchart showing the order of analysis of EMS and results that are consistent with each of 6 idealized structures |
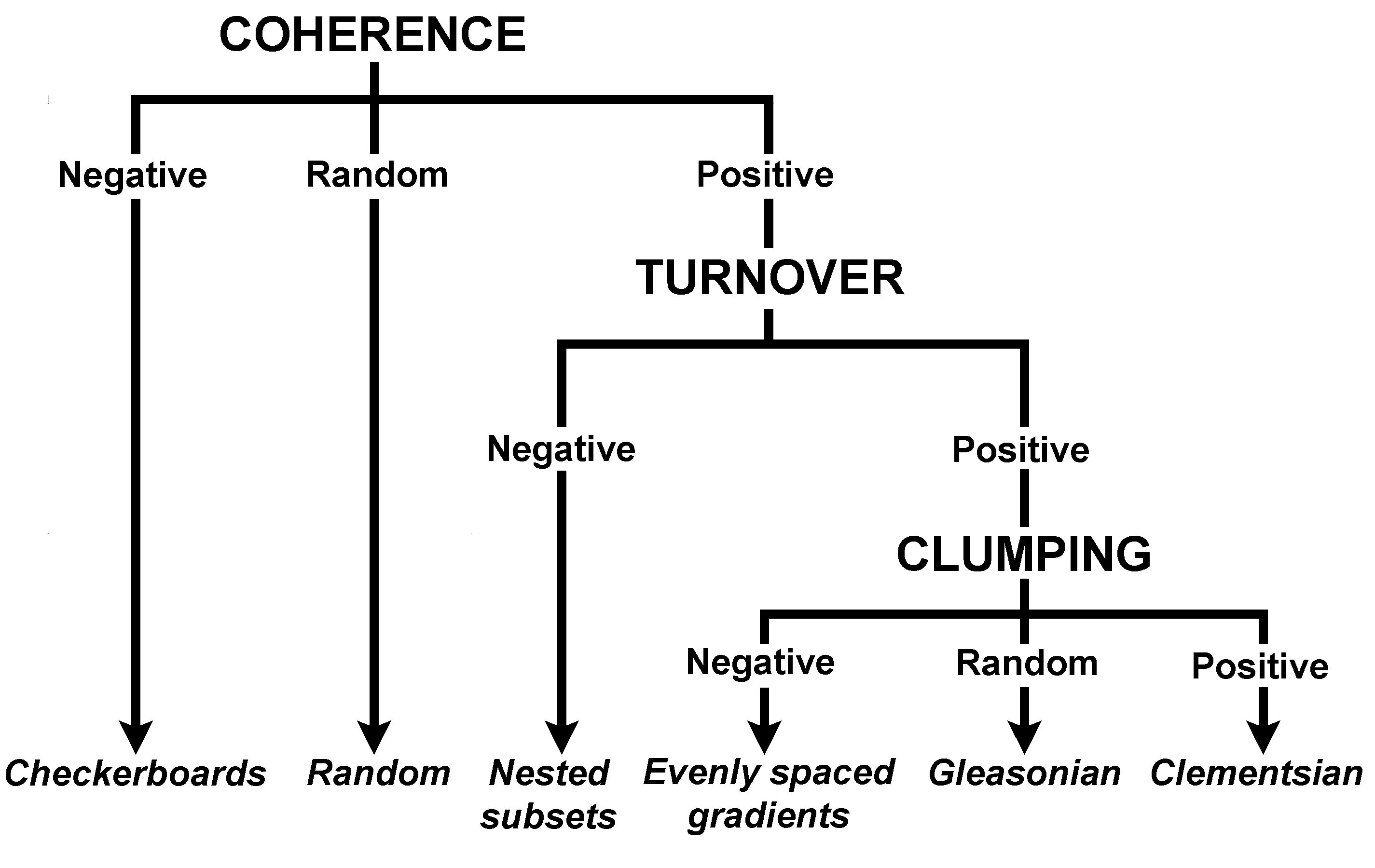
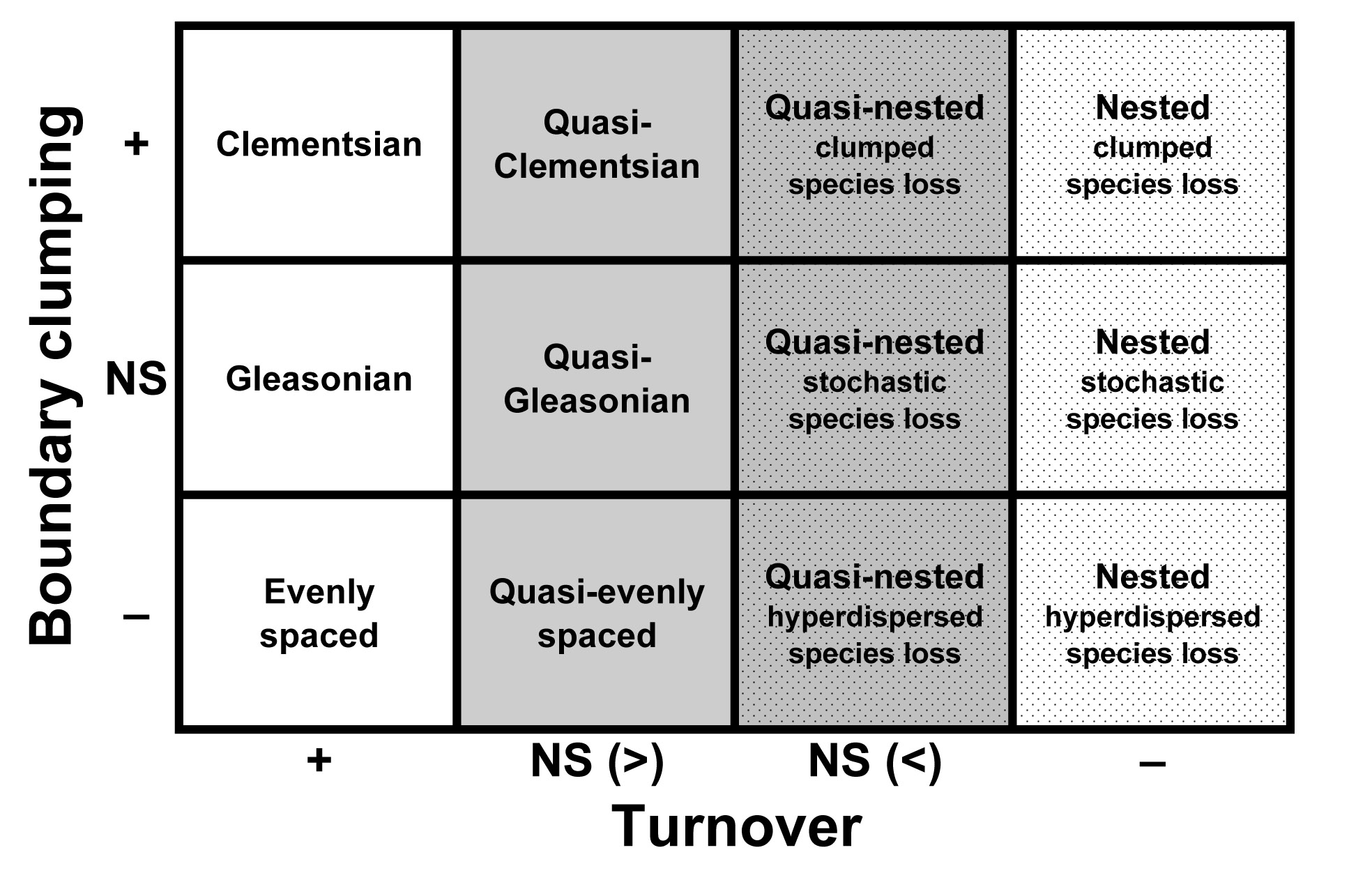
Idealized patterns of distribution.--Leibold and Mikkelson (2002) defined three elements of metacommunity structure (EMS; coherence, species turnover, and boundary clumping). In concert, analyses of these three EMS identify which idealization (checkerboard, nested, Clementsian, Gleasonian, evenly spaced and random distributions) best reflects the distribution of species along the primary axis of environmental variation as determined via reciprocal averaging (see below). Each of the non-random models of species distribution assumes that distributions of species are molded by biotic (e.g. competition, habitat associations) or abiotic (e.g. temperature, rainfall) factors that vary among sites. Although the nature of responses may differ among species, as a group the biota must respond to the same latent environmental gradient(s) for a metacommunity to evince non-random structure (i.e. coherence). If species as a group do not respond to the same environmental gradient, their distributions will not form a coherent structure, indicating that species occurrences with respect to the latent gradient are random (Leibold and Mikkelson 2002). Checkerboard distributions occur if pairs of species are mutually exclusive (Diamond 1975), and if mutually exclusive responses are independent of each other along the gradient. Distributions are nested if ranges of species that occupy a smaller portion of the environmental gradient are contained within the ranges of those that occupy a larger portion of the gradient. Distributions that exhibit turnover and whose boundaries are clumped along the environmental gradient are termed Clementsian in reference to Clements’ model of distinctive “communities” in which the boundaries of species ranges are highly coincident (Clements 1916). Distributions that exhibit turnover and whose boundaries occur idiosyncratically along the environmental gradient are termed Gleasonian in reference to Gleason’s concept of individualistic responses of species (Gleason 1926). Evenly spaced distributions exhibit turnover with boundaries that are hyperdispersed along the environmental gradient, indicating maximal differences in environmental tolerances among species.
Because all species in a community rarely respond to the same suite of environmental characteristics, empirical structures at best only approximate particular idealizations. In such cases, identification of best-fit structures is difficult, especially if objective criteria are lacking to distinguish among idealizations based on empirical data. Leibold & Mikkelson (2002) developed a set of objective criteria and a rigorous quantitative approach to simultaneously test the correspondence of an empirical structure to each of a number of hypothetical patterns of species distribution.
Quasi-structures.--The hierarchical protocol developed by Leibold and Mikkelson (2002) distinguishes among six idealized patterns, each of which has distinct conceptual foundations in ecology or biogeography (Clements 1916, Gleason 1926, Diamond 1975, Tilman 1982, Patterson and Atmar 1986). Nonetheless, combinations of coherence, turnover, and boundary clumping are possible that do not conform perfectly to any of the canonical patterns. These structures are common, including 9 of 24 coherent structures in the original publication of Leibold and Mikkelson (2002). Nonetheless, these common structures remained undefined..
We propose the recognition of four quasi-structures, each of which has a foundation that is consistent with the conceptual underpinning of Clementsian, evenly spaced, Gleasonian, or nested distributions. Each quasi-structure has the same basic characteristics as its associated idealized structure, except that range turnover is indistinguishable from random. Although some expectation of turnover is associated with non-random structures, significantly positive or negative turnover is not a fundamental characteristic of Gleasonian, evenly spaced, or nested distributions. For example, because distributions of restricted-range species are contained within those of wide-ranging species in nested structures, one would expect nested distributions to have negative turnover (i.e. fewer replacements than the average in randomly generated matrices); however, turnover need not result in statistical significance for a nested pattern to best describe a metacommunity. Similarly, some positive turnover is a fundamental character of Gleasonian or evenly spaced distributions, but statistically significant positive turnover is not required for such patterns to best describe a metacommunity. Nonetheless, the level of significance associated with turnover likely indicates the strength of structuring mechanisms, with quasi-structures resulting from weaker structuring forces than those effecting structures in which turnover is significant.
Reciprocal averaging.--Reciprocal averaging is the best indirect ordination procedure to discern sample variation in response to environmental gradients (Gauch et al. 1977). More specifically, reciprocal averaging allows composition of communities and occurrence of species to define the gradient that is important to metacommunity structure and ordinates communities and species along that gradient. Reciprocal averaging does not suffer from some of the problems associated assumptions about which variables (e.g., temperature, latitude or elevation) define species distributions, the nature (e.g., linear or curvilinear) of such relationships, or the scale of responses. Consequently, when subjected to reciprocal averaging, a metacommunity may ordinate along a gradient that integrates multiple environmental characteristics that are important to species distributions. Michael Palmer provides a brief but informative overview of ordination procedures, including reciprocal averaging.
Elements of metacommunity structure.--Metacommunity coherence has two components. First, is the idea that, all else being equal, a species distribution should span a contiguous range of an environmental gradient. In other words, one would not expect to find holes or gaps in a species fundamental niche along any particular niche dimension. In general, species are expected to exhibit coherent ranges. Second, species must exhibit coherent distributions along the same ordination axis. If this is not so, then species are not responding to the same gradient and the metacommunity does not form a coherent structure along that gradient. Groups of species that are closely related ecologically or phylogenetically tend to respond to the same environmental factors; therefore, it is expected that most well sampled metacommunities of ecologically or taxonomically similar species will exhibit coherence. Species turnover is simply a measure of the replacement of one species with another species along the gradient/ordination axis. Boundary clumping is a measure of the degree to which boundaries of species ranges are coincident (i.e., occur at the same site).
Multiple ordination axes.--Often in ordination analyses (e.g., principal component analysis) multiple axes represent biologically meaningful gradients. Reciprocal averaging “will in some cases reveal a second direction of sample variation in its second axis” (Gauch et al. 1977). In previous research on that evaluated spatial organization of Paraguayan bats in response to variation in habitat and environmental characteristics (López-González 1998, 2004; Stevens et al. 2007), analyses resulted in two axes that explained a significant amount of variation in bat species composition, with each axis representing distinct environmental gradients; although, such responses were mediated through associations of vegetation. We hypothesized that metacommunities may exhibit unique patterns of structure along multiple ordination axes and that distribution patterns on each axis may represent distinct, ecologically meaningful responses to the environment. We expanded the methodology of Leibold and Mikkelson (2002) to conduct analyses for matrices ordinated based on the second ordination axis as well as for matrices ordinated based on the first ordination axis.
 |
 |
| Brachyphylla cavernarum |
Effects of endemic species.-- The extents of most metacommunity studies, including those that employ the methods of Leibold and Mikkelson (2002), are at the local or landscape level; however, because analyses of metacommunity structure are plastic, they also can be applied in studies of biogeography. Caribbean islands exhibit the environmental variation that is necessary to evaluate the relative importance of island-specific characteristics in delimiting the geographical distribution of vagile species. Bats of the Caribbean are well studied and the distributions of species on islands are well delimited (Willig et al., 2009 and citations therein). In addition, about 30% of the bats on each island group (Bahamas, Greater Antilles, and Lesser Antilles) in the Caribbean occur on a single island in that group, which provided the opportunity to evaluate the effects of endemic species on metacommunity structure.
For an embedded absence to occur in a particular row or column of a matrix, a species must occur in at least two communities, or a community must comprise at least two species. By occurring at a single site, endemic species cannot create an embedded absence in the species row, whereas randomly generated matrices using null models without fixed row totals can form such embedded absences. The overall effect is that endemics enhance coherence. Both range boundaries for an endemic occur at the same site; therefore, endemic species may increase boundary clumping in a metacommunity. Because EMS evaluate species distributions along latent environmental gradients, and because endemic species span little (nearly none) of such gradients, understanding how their presence affects conclusions from analyses of metacommunity structure is critical.
Bahamian and Lesser Antillean bats evinced clumped range boundaries and high species turnover, consistent with a Clementsian pattern. In contrast, bat distributions in the Greater Antilles exhibited strong nestedness (Presley and Willig 2010). Removal of endemics only changed qualitative conclusions in three of 27 analyses of EMS, and affected the best-fit model in only two of nine metacommunities or metaensembles. Despite representing 1/3 of Caribbean bat species, endemics were not a primary factor affecting overall metacommunity structure.
More recently, we evaluated effects of microspatial species (those occurring at a single site) on metacommunity structure for bats, rodents, and passerine birds along an extensive elevational gradient on the eastern slopes of the Peruvian Andes. These taxa differed in proportion of species that were restricted to a single elevational band, with 31% of rodents, 21% of bats, and 10% of birds representing microspatial species. Nonetheless, removal of these species had no qualitative effect on metacommunity structure (Presley et al. submitted to GEB).
| Forms of nested subsets.--Despite the long history of analysis of nested subsets, no one has described different types of nestedness. We used analysis of boundary clumping to demonstrate that one can distinguish among 3 types of nested subsets (Presley et al. in press). These types of nestedness are discerned by the pattern of species loss as one moves from more species-rich assemblages to species-poor assemblages. Nested metacommunities may lose species in a regular fashion (hyperdispersed boundaries), at random (randomly spaced boundaries), or in groups (clumped boundaries). Nested metacommunities that lose species in an even fashion exhibit a nested substructure that is analogous to evenly-spaced distributions. Random loss of species from a nested metacommunity is analogous to individualistic responses of Gleasonian distributions, except that the individualistic responses only occur at one end of the gradient. Similarly, when groups of species are lost coincidently along the gradient; this structure is analogous to Clementsian distributions with the responses only occurring at one end of the gradient. |
|
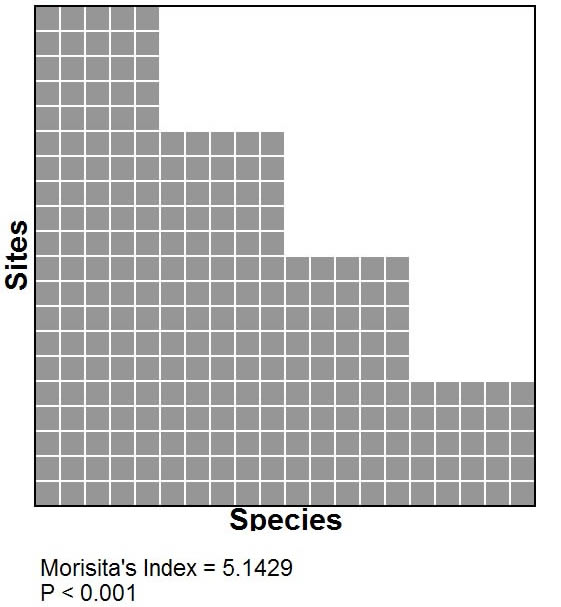
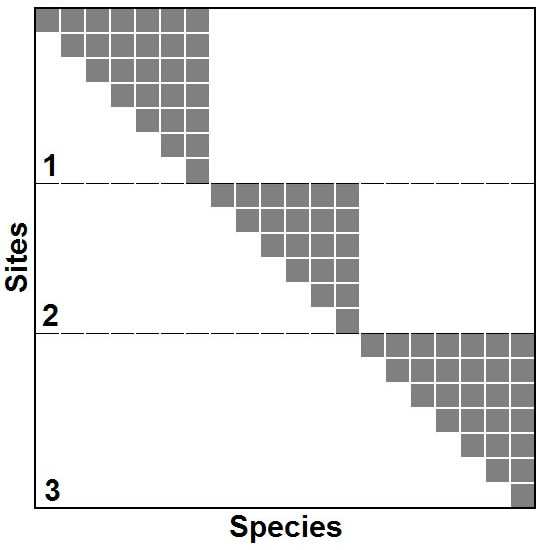
Hierarchical analyses.-- Species respond to the environment at multiple scales; therefore, distinct metacommunity structures may manifest at multiple ecologically meaningful extents. Determination of appropriate extent for analyses can be difficult, as a result researchers often use multiple arbitrarily defined spatial extents for cross-scale analyses. Metacommunities with Clementsian structure provide an opportunity to allow the biota to define the appropriate extent of a analysis while maintaining the same focus (i.e. definition of a site). For example, if a metacommunity exhibits Clementsian distributions (i.e., positive coherence, range turnover, and range boundary clumping), then it may be possible, based on the locations of clumps of boundaries, to divide a metacommunity into multiple extents based on the naturally occurring coincidence of boundaries that define Clementsian structure. Subsequently, EMS can be analyzed for each of the delineated extents, each of which may exhibit distinct structure. For example, it is possible for a group of metacommunities to each exhibit nested distributions (figure at right), but for an analysis that includes all 3 of these metacommunities to result in Clementsian structure. Each result reflects reality at a distinct scale; therefore, it is important that ecologists carefully define the spatial (or ecological) extent of their metacommunities and to carefully evaluate structure details of structure in the ordinated matrix to maximize the insights gained from analyses of EMS. |
|
|
|
| Flowchart of analyses for a comprehensive evaluation of metacommunity structure. | Coherent space showing the relationships of 12 metacommunity structures. |
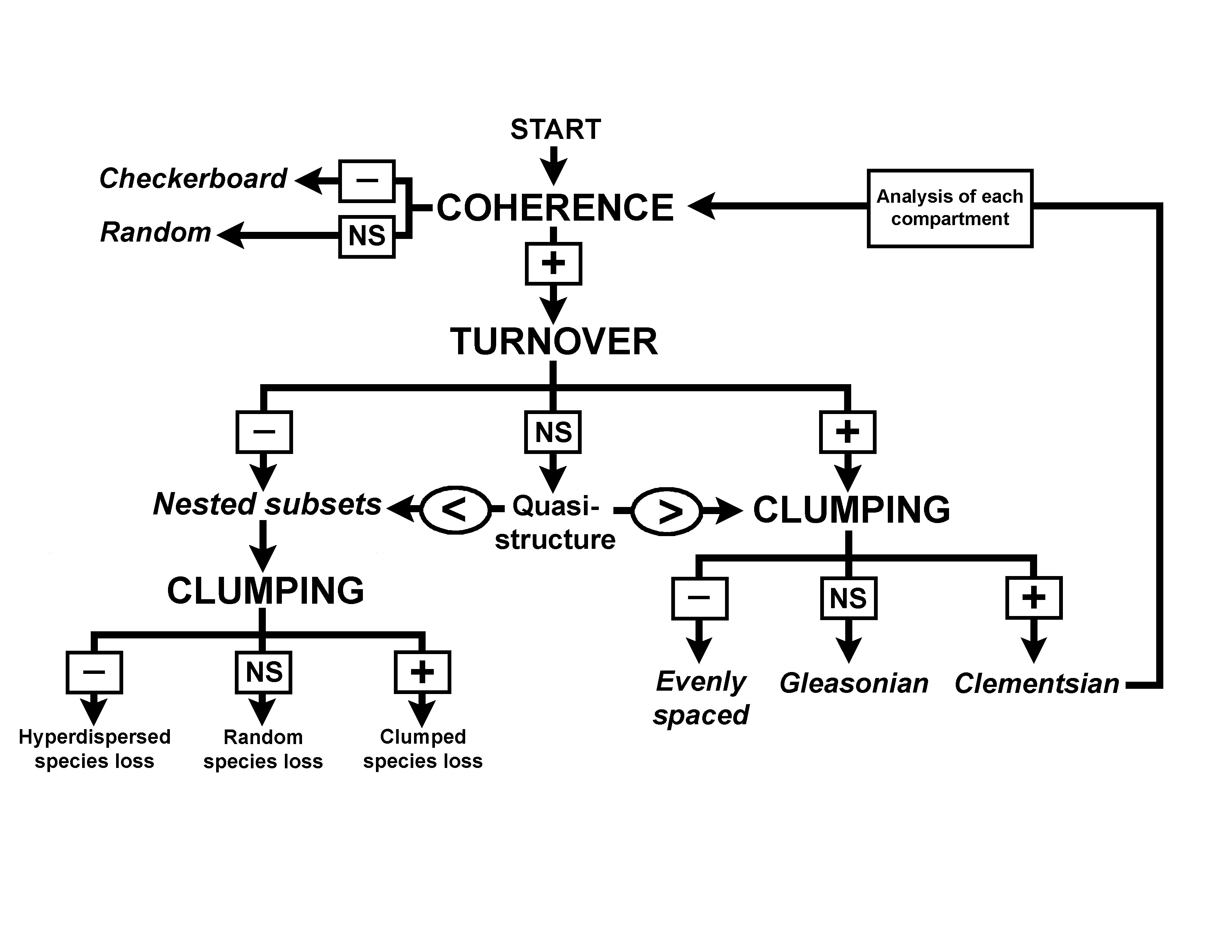
Applications.--Analysis of metacommunity structure has many uses and can be applied to many different kinds of systems. Its primary use is to evaluate patterns of species distribution along strong environmental gradients to which species are known to respond (e.g. Presley et al. 2009, 2010, 2012, Presly and Willig 2010, Willig et al. 2011, López-González et al. 2011). However, it can also be used to determine if current species distributions conform to expectations based on historical biogeographic patterns (de la Sancha et al. 2014), to determine the responses of species to anthropogenic activities at the landscape scale (Cisneros et al. submitted), and to determine the importances of host attributes in structuring parasite metacommunities (Dallas and Presley 2014).
Collaborators on Metacommunity Research
| Lab Members | Other Collaborators | ||
Related Manuscripts
Stevens, R. D., C. López-González, and S. J. Presley. 2007. Geographical ecology of Paraguayan bats: spatial integration and metacommunity structure of interacting assemblages. Journal of Animal Ecology 76:1086-1093.
Presley, S. J., C. L. Higgins, C. López-González, and R. D. Stevens. 2009. Elements of metacommunity structure of Paraguayan bats: multiple gradients require analysis of multiple axes of variation. Oecologia 160:781-793.
Presley, S. J., and M. R. Willig. 2010. Bat metacommunity structure on Caribbean islands and the role of endemics. Global Ecology and Biogeography 19:185-199.
Presley, S. J., C. L. Higgins, and M. R. Willig. 2010. A comprehensive framework for the evaluation of metacommunity structure. Oikos 119:908-917.
Presley, S. J., M. R. Willig, C. P. Bloch, I. Castro-Arellano, C. L. Higgins, and B. T. Klingbeil. 2010. A complex metacommunity structure for gastropods along an elevational gradient. Biotropica 43: 480-488.
Willig, M. R., S. J. Presley, C. P. Bloch, I. Castro-Arellano, L. M. Cisneros, C. L. Higgins, and B. T. Klingbeil. 2011. Tropical metacommunities and elevational gradients: disentangling effects of forest type from other elevational factors. Oikos 120:1497-1508.
López-González, C., S. J. Presley, A. Lozano, R. D. Stevens, and C. L. Higgins. 2012. Metacommunity analysis of Mexican bats: environmentally mediated structure in an area of high geographic and environmental complexity. Journal of Biogeography 39:177-192.
Presley, S. J., L. M. Cisneros, B. D. Patterson, and M. R. Willig . 2012. Vertebrate metacommunity structure along an extensive elevational gradient in the tropics: a comparison of bats, rodents, and birds. Global Ecology and Biogeography 21:968-976.
Willig, M. R., S. J. Presley, C. P. Bloch, and J. Alvarez. 2013. Population, community, and metacommunity dynamics of terrestrial gastropods in the Luquillo Mountains: a gradient perspective. In: Ecological Gradient Analyses in a Tropical Landscape, G. González, M.R. Willig, and R. B. Waide, editors. Ecological Bulletins 54:117-140.
Dallas, T., and S.J. Presley. 2014. Relative importance of host environment, transmission potential, and host phylogeny to the structure of parasite metacommunities. Oikos 123: 866-874.
de la Sancha, N., C.L. Higgins, S.J. Presley, and R.E. Strauss. 2014. Metacommunity structure in a highly fragmented forest: has deforestation in the Atlantic Forest altered historic biogeographic patterns? Diversity and Distributions 20: 1058-1070.
Cisneros, L.M., M.E. Fagan, & M.R. Willig. 2014. Season-specific and guild-specific effects of anthropogenic landscape modification on metacommunity structure of tropical bats. Journal of Animal Ecology 84:373-385.
Bloch, C.P. and B.T. Klingbeil. 2015. Anthropogenic factors and habitat complexity influence biodiversity but wave exposure drives species turnover of a subtropical rocky inter-tidal metacommunity. Marine Ecology doi: 10.1111/maec.12250.
References
Clements, F. E. 1916. Plant succession: an analysis of the development of vegetation. Carnegie Institution of Washington, Washington, DC.
Diamond, J. M. 1975. Assembly of species communities. Ecology and evolution of communities (ed. by M.L. Cody and J.M. Diamond), pp. 342-444. Harvard University Press, Cambridge, Massachusetts.
Gauch, H. G., R. H. Whittaker, and T. R. Wentworth. 1977. A comparative study of reciprocal averaging and other ordination techniques. Journal of Ecology 65:157-174.
Gleason, H. A. 1926. The individualistic concept of the plant association. Bulletin of the Torrey Botanical Club 53:7-26.
Holyoak, M., R. D. Holt, and M. A. Leibold. (eds). 2005 Metacommunities: spatial dynamics and ecological communities. University of Chicago Press, Chicago, Illinois.
Hoagland, B.W. and S. L. Collins. 1997. Gradient models, gradient analysis, and hierarchical structure in plant communities. Oikos 78:23-30.
Hofer, U., L. F. Bersier, and D. Borcard. 1999. Spatial organization of a herpetofauna on an elevational gradient revealed by null model tests. Ecology 80:976-988.
Hausdorf, B. and C. Hennig. 2007. Null model tests of clustering of species, negative co-occurrence patterns and nestedness in meta-communities. Oikos 116:818-828.
Leibold, M.A., M. Holyoak, M. Mouquet, P. Amarasekare, J. M. Chase, M. F. Hoopes, R. D. Holt, J. B. Shurin, R. Law, D. Tilman, M. Loreau, and A.Gonzalez. 2004. The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters 7:601-613.
Leibold, M.A. and T. E. Miller. 2004. From metapopulations to metacommunities. Ecology, Genetics and Evolution of Metacommunities (ed. by I. A. Hanski and O. E. Gaggiotti), pp. 133-150. Elsevier Academic Press, Burlington, Massachusetts.
Leibold, M.A. and G.M. Mikkelson. 2002 Coherence, species turnover, and boundary clumping: elements of meta-community structure. Oikos 97:237-250.
López-González C. 1998. Systematics and zoogeography of the bats of Paraguay. Unpublished PhD. Dissertation, Texas Tech University, Lubbock, TX.
López-González C. 2004 Ecological Zoogeography of the bats of Paraguay. Journal of Biogeography 31:33-45.
Willig, M. R., S. J. Presley, C. P. Bloch, and H. H. Genoways. 2009. Macroecology of Caribbean bats: effects of area, elevation, latitude, and hurricane-induced disturbance. In: Island bats: evolution, ecology, and conservation (T. Fleming, and P. Racey, Eds.). University of Chicago Press, pages 216-264.