Stenoderma
rufum Desmarest, 1820 |
||
Red Fig-eating Bat or Murciélago Frutero NativoM.R. Gannon, M. Rodríguez-Durán, A. Kurta, and M.R Willig Taxonomy
The genus Stenoderma (Koopman, 1993) is placed in the subfamily Stenodermatinae, along with Artibeus and 16 other genera. However, unlike two living Artibeus, Stenoderma contains only a single species, Stenoderma rufum, which includes 2 subspecies, S. r. rufum and S. r. darioi. The type specimen for S. r. rufum, and, hence, for the species, was taken at an unknown locality in the early 1800’s, but given the present distribution of subspecies and the appearance of the type specimen, biologists believe that the original specimen was from the Virgin Islands. The description of S. r. darioi is based on a bat captured in 1965, near El Yunque Peak, in eastern Puerto Rico (Hall and Tamsitt, 1968). NameThe word Stenoderma comes from the Greek words stenos and derma which mean "narrow” and “skin," respectively, and the two words most likely refer to the narrow tail membrane of this species. The specific epithet rufum is Latin for red and refers to the occasionally reddish brown color of this animal on the Virgin Islands, where the type presumably was collected. The subspecific name darioi is in honor of Darío Valdivieso, a neotropical biologist who published a number of papers concerning bats in the 1960s and 1970s. The history of the bat named Stenoderma rufum is rather intriguing and shows the confusion that can result when museum specimens, particularly type specimens, are not cared for properly (Anthony, 1925). Although A. Demarest was the first to use the current scientific name in 1820, the animal actually was introduced to the scientific world a few years earlier as “le Sténoderme roux” by Etienne Geoffroy Saint-Hilaire, a prolific French taxonomist of the early 19th century. Geoffroy Saint-Hilaire (1818) published a description of the species based on the skin and skull of a specimen housed in the National Museum of Natural History in Paris. However, the specimen apparently had been mislabeled, and Geoffroy Saint-Hilaire reported that it was from Africa. In 1869, W. Peters attempted to examine the holotype, but he found only the skin; the skull apparently had disappeared since the time of Geoffroy Saint-Hilaire. A few years later, Peters went to the same museum and examined a different specimen, this time consisting of only a skull, which had been given the name Artibeus unduatus by another French taxonomist, P. Gervais. Peters noted that the skull of A. unduatus appeared identical to the one that Geoffroy Saint-Hilaire had drawn for his published description of “le Sténoderme roux.” Peters surmised that the skull of A. unduatus was actually the long-lost skull of S. rufum, and his conclusion generally is accepted today. For almost a century, the red fig-eating bat was known only from that single, poorly preserved specimen in the Paris Museum. Over the years, biologists had commented that the specimen’s anatomical traits strongly indicated that it represented a species from the New World and not from Africa, but proof was lacking. Finally, in 1916, fossilized remains of this species were found in caves on Puerto Rico, thus confirming its presence in the Western Hemisphere (Anthony, 1918). Nevertheless, whether the species was still living or had become extinct since its original description was not settled until 1957, when James Bee captured three individuals on St. John, in the Virgin Islands (Hall and Bee, 1960). Distribution and StatusThe red fig-eating bat is endemic to the Greater Antilles and occurs on very few islands. S. r. rufum is known from the Virgin Islands of St. John and St. Thomas, although it is quite rare there and we are not aware of any specimens taken on the Virgin Islands in the last 30 years. S. r. darioi was thought to live only on Puerto Rico, but a juvenile red fig-eating bat recently was captured on Vieques, suggesting the presence of an additional population on that island (Rodríguez-Durán, 2002). This bat appears most common in the Luquillo Experimental Forest, where, at one time, this species represented at least 25% of bats captured in tabonuco forest (Willig and Bauman, 1984). Nevertheless, only about 50 individuals have been taken in other parts of Puerto Rico and on Vieques (Gannon, 2002), and whether the paucity of records away from Luquillo represents a habitat preference by the species or simply lack of field work in habitats other than tabonuco forest is unknown.
The International Union for Conservation of Nature and Natural Resources (IUCN) classifies the red fig-eating bat as “vulnerable” (category “VU a1c”) to extinction (Hutson et al., 2001). This designation indicates that, although not critically endangered at present, the species may be facing a high risk of extinction in the medium-term future. The red fig-eating bat has the smallest geographic distribution of any species of bat that occurs on Puerto Rico and one of the smallest in the Antilles. The IUCN declared the species vulnerable because of a suspected decrease in the already small geographic distribution of this island endemic and because of the devastating effects of hurricanes Hugo and Georges (detailed below). Measurements and Dental Formula Total length is 60–69 mm; length of hind foot, 12–18 mm; height of ear, 15–20 mm; length of forearm, 48–52 mm; body mass is 20–31 g. As in many species of bat, female red fig-eating bats are marginally, but consistently, larger than males in many traits (Jones et al., 1971; Gannon et al., 1992). Dental formula is incisors 2/2, canines1/1, premolars 2/2, molars 3/3 = 32.
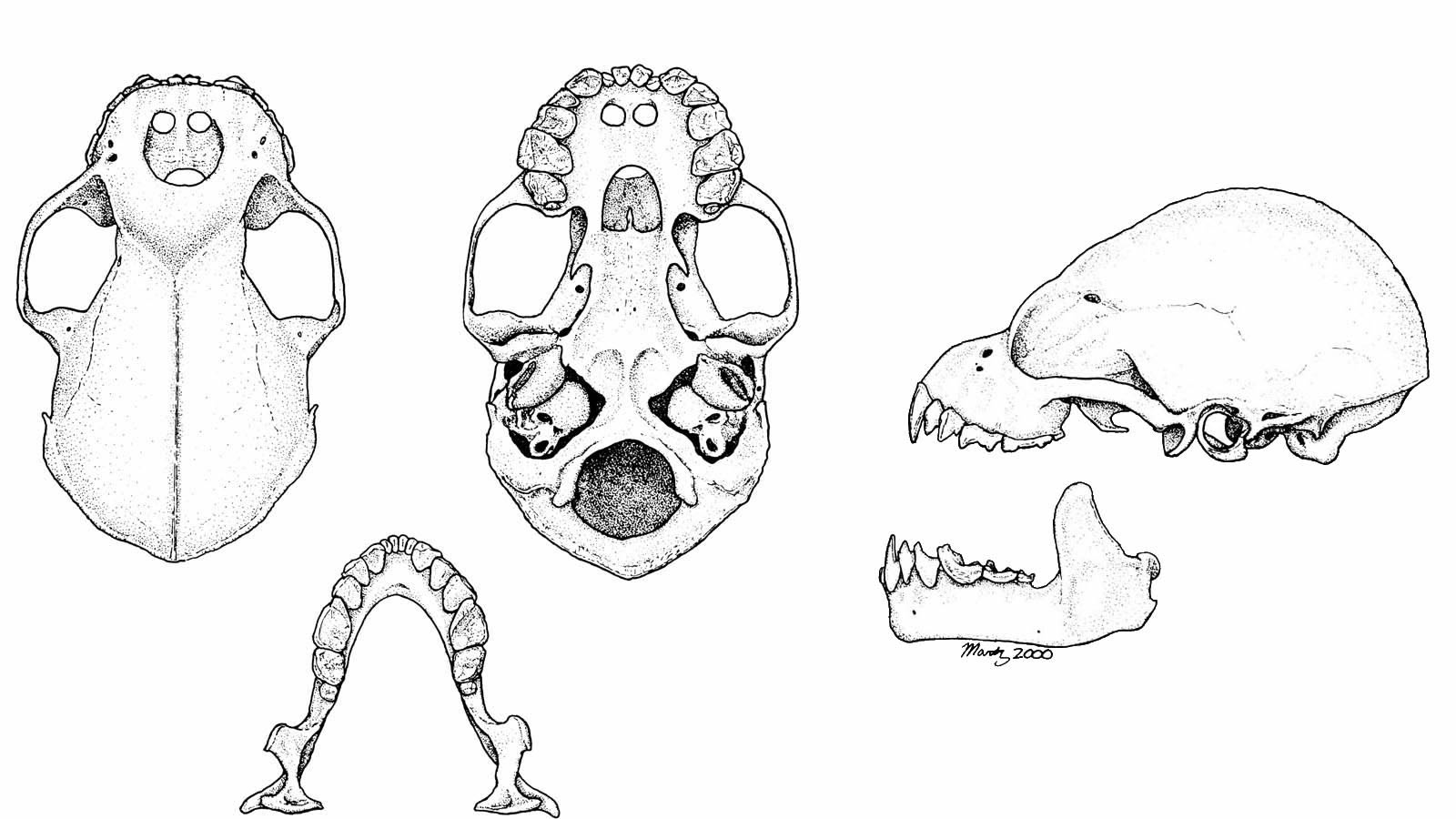
DescriptionAlthough red fig-eating bats from the Virgin Islands may have a reddish tint to their fur, those from Puerto Rico typically appear brown or tan. Ventral hairs are tipped with gray and appear somewhat lighter. A dab of white occurs on the side of the head below each ear, and both sexes also have a white shoulder patch that is about 4 mm in diameter (Figs. 9 and 44). Underneath the shoulder patches of adult males, there are glands that emit a strong musk-like odor. A tail is absent, and the narrow interfemoral membrane has only a sparse covering of hairs. A well-developed nose-leaf easily establishes this species as a member of the Phyllostomidae, and the bat’s gentle disposition, combined with medium body size, white patches of fur, and lack of a tail, distinguish this animal from all other phyllostomids on Puerto Rico. A lack of W-shaped ridges on the cheek teeth and presence of incisive foramina in the palate indicate that the skull belongs to one of the phyllostomids of Puerto Rico (Fig. 45). The rostrum of a red fig-eating bat differs from that of other phyllostomids on the island in that it is obviously wider than it is long. Skulls of the Jamaican and Antillean fruit bats are moderately similar in construction, but their rostrums are not as short and the greatest length of their skulls (more than 26 millimeters) exceeds that of a red fig-eating bat (21–24 millimeters).
Natural HistoryOnly the population in the Luquillo Mountains has been studied extensively, and almost all that is known of the natural history of this phyllostomid comes from animals living there. The red fig-eating bat, like other members of its subfamily, is primarily a frugivore, but despite its common name, there is no evidence that it actually eats figs. In the Luquillo Mountains, the most commonly eaten fruits are from the trumpet tree, bullet-wood, and sierra palm, with a few bats taking elder as well (Genoways and Baker, 1972; Scogin, 1982; Willig and Bauman, 1984; Willig and Gannon, 1996). Frugivorous bats play an important role as seed dispersers in tropical ecosystems, and the red fig-eating bat may be the major, and perhaps only, disperser for bullet-wood in the tabonuco forest of Puerto Rico (You, 1991). Foraging occurs both above and below the forest canopy (Jones et al., 1971). Daily movements of the red fig-eating bat have been documented extensively in tabonuco forest through radio-tracking (Gannon, 1991; Gannon and Willig, 1994a). Unlike other phyllostomids that live in large colonies in caves, both male and female red fig-eating bats are solitary and roost among the leaves of the forest canopy. These bats frequently change their roosting location, and the same site seldom is occupied more than once. Home range is small, only about 2.5 hectares on average. Subadults have larger home ranges than do mature adults, perhaps because young animals are less experienced at finding food or possibly because adults exclude them from trees with the best quality or greatest quantity of fruit. In any event, there is no evidence of territoriality, and individual home ranges overlap considerably. The small home range indicates that the red fig-eating bat spends little time or energy commuting to feeding areas. This behavior contrasts markedly with that of other fruit-eating species, such as the Jamaican fruit bat, that roost in colonies and often travel great distances to foraging grounds (Morrison, 1978a, 1978b). Red fig-eating bats are active throughout the night and return to the day roost just before dawn. Although some species of bat avoid flying in bright moonlight, presumably to avoid predation (Morrison, 1978c), the red fig-eating bat, like the Jamaican fruit-eating bat, does not alter its behavior in response to changing lunar illumination on Puerto Rico (Gannon and Willig, 1997). This lack of a response to moonlight may be related to the fact that the red fig-eating bat evolved on isolated islands with no visually oriented, nocturnal predators capable of capturing this phyllostomid. Hurricane Hugo passed over the Luquillo Experimental Forest in September 1989 and caused large-scale damage and defoliation (Walker, 1991; Walker et al., 1996). Immediately after the hurricane, captures of the red fig-eating bat decreased by 80%, and population levels did not return to pre-hurricane levels until 5 years following the storm (Gannon, 2002). Although juveniles made up 30–40% of the catch before Hurricane Hugo, this proportion fell to 10–20% after the storm, demonstrating an effect on reproductive success as well. Hugo greatly reduced availability of fruits eaten by this species, and bats that remained in the forest responded by increasing size of their home range by a factor of five. Although number of red fig-eating bats greatly declined, this species still was the most abundant frugivorous bat in tabonuco forest during the first year after the hurricane (Gannon and Willig, 1994b), and consequently, the red fig-eating bat probably played an important role in disseminating seeds of early successional plants following the storm. This foliage-roosting species was dealt another severe blow by Hurricane Georges in 1998, and the population in the Luquillo Experimental Forest showed no sign of recovery over 4 years later (Gannon, 2002). Pregnant females have been captured on Puerto Rico in January, March, June, July, and August, and lactating bats are known from March, May, June, and July. Males with descended testes have been taken in January, February, March, June, July, and August. As with many other species of bat on Puerto Rico, no data on the reproduction of red fig-eating bats are available for September through December. Females often are simultaneously pregnant and lactating, which indicates that females likely undergo a postpartum estrus and that the species is polyestrous (Genoways and Baker, 1972; Willig and Bauman, 1984; Gannon and Willig, 1992). Nonetheless, births do not appear restricted to particular times of year, i.e., red fig-eating bats exhibit asynchronous polyestry. This reproductive pattern is characteristic of species that live in areas where the abundance of food does not vary greatly on a seasonal basis (Wilson, 1973, 1979), and this appears true in the tabonuco forest of the Luquillo Mountains (Devoe, 1990). Among mammals, bats have the largest offspring, relative to the size of the mother, and among bats, phyllostomids have some of the largest neonates (Kurta and Kunz, 1987; Hayssen and Kunz, 1996). In the red fig-eating bat, the single newborn weighs about 37% of the mother’s body mass after she has given birth, and its forearm already is 59% as long as that of its mother (Tamsitt and Valdivieso, 1966). Young of this species are born with dense, grayish hairs covering the back, and the white shoulder patch is obvious at birth. The face, in contrast, is mostly naked and appears pink. Eyes are open at birth. Females give birth in the usual resting position with the head facing down; youngsters are delivered headfirst and received by the mother in cupped wings. The only ectoparasites reported for the red fig-eating bat are four species of mite (Gannon and Willig, 1994b, 1995), including one spinturnicid (Periglischrus iheringi) and three labidocarpids (Paralabidocarpus artibei, P. foxi, and P. stenodermi). The wing mite P. iheringi is the most common ectoparasite and occurs on 35% of red fig-eating bats, with infested bats harboring about four mites each on average. As in the Jamaican fruit bat, juveniles support a greater number of parasites than do adults, perhaps because adults are more efficient at grooming and removing the mites. Levels of infestation are low compared with other Puerto Rican species of bat that have been studied, and this may be related to the solitary roosting habit of the red fig-eating bat. Roosting alone would limit opportunities to obtain parasites by direct contact with other individuals, and this solitary behavior also would minimize the possibility of uninfected animals directly contacting parasites, such as argasid ticks, that spend part of their life cycle off the host and in the roost area. |


Please contact steven.presley@uconn.edu or michael.willig@uconn.edu if you have questions, comments, or corrections. Unless otherwise indicated, figures, tables, and images that appear on this page may not be used, reproduced, copied, or posted elsewhere, for any reasons, without permission. |